Marine Ecology Research
Opportunities
Dr. Walters’ Lab
Dr. Walters’ Lab
Recent
papers by Bertness and Silliman raised the profile of intertidal marsh snails
and posed many intriguing questions about the role of snails in salt marsh
dynamics. Along with Dr. Hutchens, CCU
Biology, we are examining the following questions focusing on the marsh
periwinkle, Littoraria irrorata, as a model organism:
1) What are the natural movement patterns of
snails and what factors affect those movements?
2) What are the long-term contributions of
snails to salt marsh ecosystem dynamics and energy flow?
3) Do L.
irrorata and S. alterniflora interact in the marsh (e.g., snails gardening plant fungi, plants
developing physical/chemical defenses to snail foraging, etc.)?
Coastal
industrial, residential and recreational development has accelerated the loss
of critical estuarine habitats (e.g., salt marshes, oyster reefs). In collaboration with Dr. Coen, SCDNR, we
have been examining the possibility of establishing structurally and
functionally similar marsh and reef habitats in GA and SC. We also have been assessing the most
effective methods and costs/benefits of restoration efforts.
Crustacean Dynamics
Harpacticoid
copepods (far right) are the first or second most numerous marine
micrometazoans living in or on marsh sediments and plants. Copepod communities associated with Spartina
stems are isolated from one another at least 2x per day. We have been studying the population and
community dynamics of the stem-copepod association in order to examine the
assumptions and predictions of Metapopulation Theory.
Habitat
Boundaries
Increasing
residential and industrial development of coastal environments (top image,
right) has resulted in increased loss of natural terrestrial-saltmarsh
boundaries (bottom image, right). With
Drs. Hutchens, Luken and Koepfler, CCU Biology & Marine Science, we have
been examining the boundary loss effects on the ecological dynamics of
high-marsh communities.
The above are just a
few examples of ongoing research projects in which students and I have been
involved over the past few years. If
you are interested in participating in any of these or other research projects
please contact me at kwalt@coastal.edu, 349-2477 or stop by my office in SCX4.



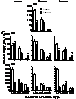




Blue
crabs are reported to exert top-down regulation on snail numbers in the marsh
limiting the snail’s ability to over-graze cordgrass. Note the numbered tags on the snails
enabling mark-recapture studies.
Salinity
differences between Murrells Inlet
barrier island and mainland developed and undeveloped sites. Salinities were greater within developed
site sediments.
Movement
of marked snails out of 0.25 m2 plots in which 0 or 50% of the Spartina
stems had been removed. Inconsistent
with past studies, data suggest that snails are very mobile within salt marsh
habitats.

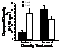

Snail
density differences between Murrells
Inlet barrier island and mainland developed and undeveloped sites. Littoraria were only present in
significant numbers in developed sites whereas Melampis densities were
greater in undeveloped sites.
Murrells
Inlet developed (top) and undeveloped (bottom) mainland sites.
Salt
marsh restoration project (GA) showing initial Spartina planting (top
left) and restored (top right) and natural marsh (bottom) growth 2 yrs. later.
Ongoing
oyster reef restoration experiment in Murrells Inlet testing the effectiveness
of bag shells and stakes as oyster recruitment and reef restoration methods.
Micrometazoan
density differences on Spartina standing-dead stems collected during
low and high tides in a GA saltmarsh.
Greater copepod numbers are found on stems during high tides.
Examples
of male, female and copepodite harpacticoid copepods.

Harpacticoid
copepod numbers trapped in the water column of a GA salt marsh during day and
night incoming and outgoing tides. Diel
and tidal differences suggest copepods are dispersing actively through the
water column.
Copepod
numbers on standing-dead Spartina stems maintained within experimental
chambers in which initial densities were either unchanged or increased. Day 0 to Day 5 changes indicated copepod
migrations between stems.
Seasonal
changes in L. irrorata densities among low-, mid- and high-marsh zones
within the Waites Island barrier salt marsh.
Distributional differences may result from differences in settlement or
predation pressure.
New and
old shoot leaf differences in S. alterniflora cinnamic acid composition
within high- and low-marsh zones.
Cinnamic acid concentrations consistently were greater in old stem
leaves and may inhibit L. irrorata feeding.
Yearly
structural differences between natural and restored marsh S. alterniflora live,
standing-dead and new shoot densities within GA high-, mid- and low-marsh
zones. Stem densities were similar
between natural and restored marshes by 1998, ≈2 yrs. after restoration.

Yearly
differences in Gukensia demissa, ribbed mussel, shell and tissue
growth between natural and restored GA
salt marshes. Except for the low-marsh,
mussel growth was greater within the natural marsh and indicated continued
functional differences within the restored salt marsh.


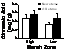



(Last updated
1/04)